Research Overview

|
The Siders lab leverages diverse quantitative approaches from across ecology and fisheries science to aid natural resource management, generate robust ecological inference, and develop new analytical approaches. We use the challenges of data deficiency and big data to drive innovation, integration, and collaboration in our research.
|

|
FISHERIES MANAGEMENT


Swordfish for sale at the Honolulu Fish Auction

Opah for sale at the Honolulu Fish Auction
|
Protected Species Population Assessment — As part of a collaboration with the NOAA Pacific Islands Fisheries Science Center, NOAA Pacific Islands Regional Office, and the Western Pacific Regional Fisheries Management Council, Dr. Siders models the trajectory of two populations of protected sea turtle species: western Pacific Leatherbacks and north Pacific Loggerheads. This species are caught incidentally on the Hawai'i and American Samoa pelagic longline fisheries. Using a Bayesian population trend estimation and a "take" model with demographic and fishery stochasticity, the team estimated negligible impacts of the Hawai'i shallow-set, deep-set, and the American Samoa longline fishery on the two populations. Unfortunately, other external threats are negatively impacting the trend of western Pacific Leatherbacks, which are likely to go extinct before the end of the century. Our findings are published in two NOAA Technical Memoranda: TM-PIFSC-95 & TM-PIFSC-101 covering the Hawaii shallow-set pelagic longline fishery for Swordfish, the Hawaii deep-set pelagic longline fishery for Bigeye Tuna, and the American Samoa pelagic longline fishery for Albacore and Yellowfin Tuna. This is an active area of research that supports Biological Opinions and Section 7 consultations under the Endangered Species Act, discussions of spatial management and best practices at the WPRFMC, and contributes to intergovernmental management of protected species in the Pacific.
|
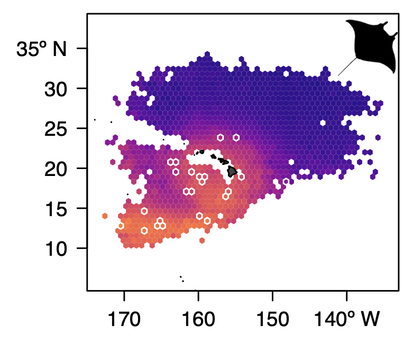 Ensemble Random Forest predictions of Giant Manta Ray interactions around Hawaii
Ensemble Random Forest predictions of Giant Manta Ray interactions around Hawaii
Ecosystem-based Fisheries Management — The Siders' lab has ongoing research building EBFM and dynamic ocean management tools for the Central North Pacific in collaboration with the NOAA Pacific Island Fisheries Science Center and Western Pacific Regional Fisheries Management Council. The two primary pelagic fisheries are the Hawaii-based Bigeye Tuna and Swordfish using longlines. Protected sea turtle, cetacean, seabird, and shark species are infrequently encountered by the fisheries but, due to their conservation status, each interaction is a potential cause for concern. But with the moving target of pelagic species, it can be difficult to effectively maintain sustainable fisheries alongside protected pelagic species. Dynamic Ocean Management seeks to solve this by using risk maps or dynamically closed spatial areas to reduce bycatch. Current research is exploring the effects on bycatch rates by building dynamic ocean management products then using management strategy evaluation to simulate how fishers might respond and evaluate the performance of the tools. An example of this work is an evaluation of the long-term dynamic informational product, TurtleWatch, published in Biological Conservation. Most current work is built on the backbone of Ensemble Random Forests, a machine-learning algorithm implementation, that was developed to support the estimation of the distribution of rare bycatch interactions. One example of these applications is a hindcast assessment of dynamic spatial zones for potentially reducing western Pacific leatherback bycatch with New Zealand longline fisheries. We've also pursued using the Ensemble Random Forests tool for developing machine-learning methods for estimating the amount of bycatch and found they work really well except for hyper-rare bycatch.
This is an active area of research with funding from the NOAA Pacific Islands Fisheries Science Center to develop protected species management and EBFM tools and applications for the US-flagged central Pacific fisheries.
BAYESISAN
APPROACHES
FOR
LIFE HISTORY
APPROACHES
FOR
LIFE HISTORY
Life history is a critical component of a species' ecology involving many physiological and biological processes such as growth, maturity, and reproduction. For many species, information on life history can be hard to acquire but essentially for determining the risk to a species' survival from detrimental actions. Our lab works to generate Bayesian approaches to integrate various data sources on life history for a wide-variety of species. Below are published examples of this research.
|
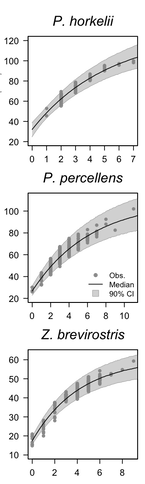
Brazilian Guitarfish — Led by Dr. Fabio Caltabellotta, Dr. Siders developed a Bayesian age-growth model for Pseudobatos horkelii, Pseudobatos percellens, and Zapteryx brevirostris in southern Brazil. The resulting model was used to estimate age and growth parameters for these three priority guitarfish species listed as Critically Endangered, Near Threatened, and Vulnerable by the International Union for the Conservation of Nature. Now published: doi: 10.1111/jfb.14123

(Above) Bowtie sections of guitarfish vertebrae for the three Brazilian guitarfish species showing off the classic banding patterns left behind by annual growth spurts. (Left) Age-growth relationships for three Brazilian Guitarfish species. Age is on the x-axis in units of years and growth is on the y-axis in units of centimeters of total length.
|
|
Brazilian Electric Ray — Led by Dr. Fernanda Rolim, a postdoc at Universidade Estadual Paulista in São Paolo, Brazil, Dr. Siders extended the Brazilian guitarfish model to estimate two-dimensional growth of Narcine brasiliensis, the Brazilian Electric Ray, in southern Brazil. The joint estimation of length-weight and age-growth parameters was added along with derivations of age at maturity, longevity, and mortality at age. A significant component was incorporating uncertainty in size at birth into the von Bertalanffy growth model.
Now published: doi: 10.1111/jfb.14378 |
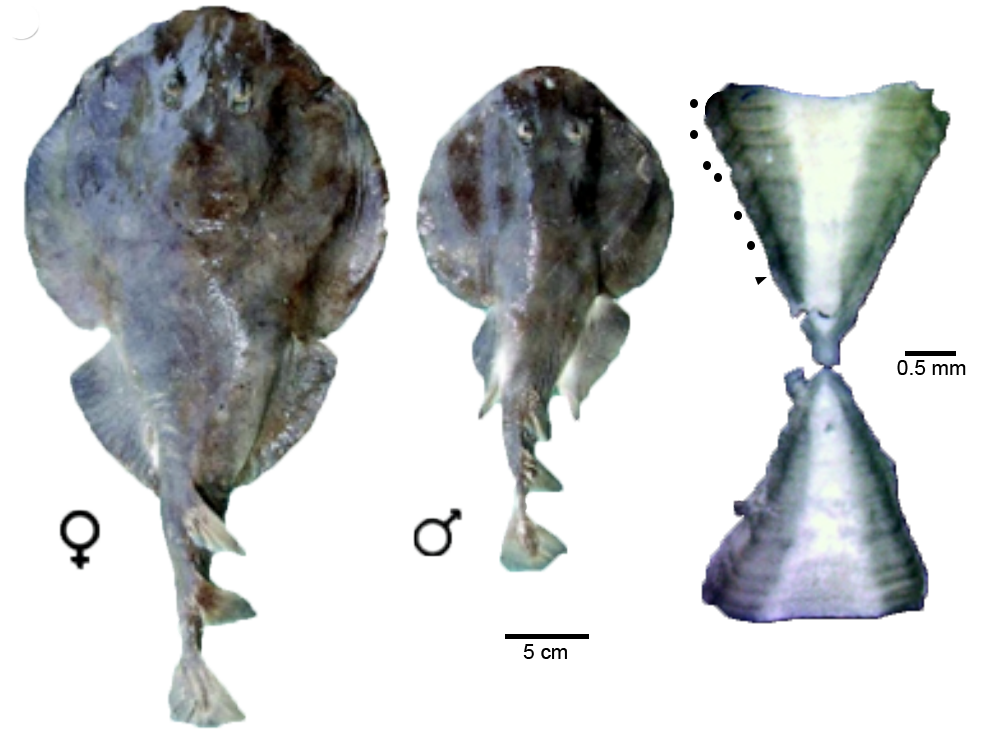
Female (left) and male (center) Brazilian Electric Ray with their vertebral section (right) used to age individuals
|

(Above) Silhouettes of Goblin Sharks through the ages. (top) 1898 in the description of the species by David Starr Jordan. (top middle) 1904 by King Bragança of Portugal. (top bottom) 1909 by Hussakof and the renaming of Mitsukurina to Scapanorhynchus for a time. (bottom) 1981 by Cadenat and Blache in Requins de Méditerranée et d' Atlantique.
|
Goblin Shark — Led by Dr. Siders and Dr. Fabio Caltabellotta, we estimated the first age-growth relationship for Goblin Sharks (Mitsukurina owstoni), one of the largest deepwater sharks. Fabio developed a method to highlight the classic banding pattern that allowed an age reading on a specimen caught in Brazil in 2008. I developed a Bayesian age-growth model that used back-calculated lengths at age from this specimen, data on maximum male sizes, and data on size at birth to estimate the first age-growth parameters for the species. We are excited to age more specimens in the future. Now published: doi.org/10.1071/MF19370
|
 (Above) Vermilion Snapper otolith
(Above) Vermilion Snapper otolith
Vermilion Snapper — Led by Dr. Derek Chamberlin and Dr. Will Patterson, we validated the age of Vermilion Snapper (Rhomboplites aurorubens), an important species to Gulf of Mexico reef fish fisheries, using a bomb radiocarbon chronometer. We found the ageing of Vermilion Snapper to be accurate, the oldest fish in our samples was 28 years old. But, we also found the species to exhibit highly variable length-at-age with little differences between sexes, estimated with a Bayesian regression. This has important consequences to fisheries stock assessments, which often use age-length keys to turn large samples of fish lengths to ages. Now published: doi.org/10.1016/j.fishres.2023.106732
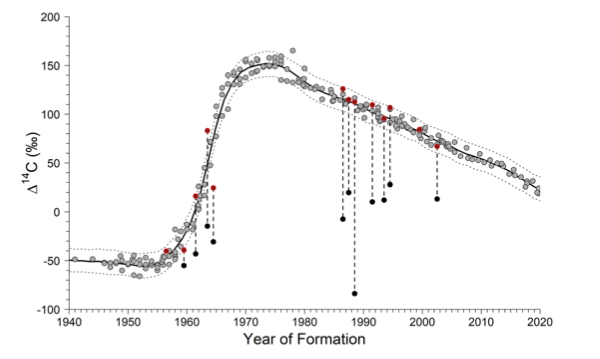 (Above) Bomb radiocarbon chronometer for the Gulf of Mexico (black line) with eye lens samples (red circles) and otolith samples (black circles) of Blackbelly Rosefish
(Above) Bomb radiocarbon chronometer for the Gulf of Mexico (black line) with eye lens samples (red circles) and otolith samples (black circles) of Blackbelly Rosefish
Blackbelly Rosefish — Led by Dr. Derek Chamberlin and Dr. Will Patterson, we validated the ageing of and estimated the growth of Blackbelly Rosefish (Helicolenus dactylopterus) in the northern Gulf of Mexico. Dr. Siders led the development of a Bayesian penalized spline model to calculate the bomb radiocarbon chronometer and estimate the accuracy of the age observations. Together, Drs. Siders and Chamberlin used the validated ages to extend the von Bertalanffy growth model used for other species to account for inter-reader error in this incredibly long-lived fish, the oldest sample being 98 years old! Most importantly, this paper showed that the use of eye lens to get birth year estimates was valid and overcame the limits of otoliths, which tend to not work for deepwater fishes. Now published: doi.org/10.1038/s41598-023-34680-0
River Cooters — Led by Eric Munscher through the Turtle Survival Alliance North American Freshwater Turtle Research Group, 20 years of mark-recapture sampling was conducted on four Pseudemys species (P. c. suwanniensis, P. nelsoni, P. peninsularis, P. texana) at five freshwater springs, four in Florida and one in Texas. Dr. Siders led the development of a hierarchical Bayesian von Bertalanffy mark-recapture growth model to separate out the species-specific growth from individual variation and influences of the environment on growth across all species and sites simultaneously. Growth estimates were the first for three of the taxa, first mark-recapture for four of the taxa, the sampling found larger specimens than ever recorded for all of the taxa, and the length at maturity estimates matched well previous studies indicating that life-history invariants are a viable strategy for estimating age at maturity in freshwater turtles. In all, this publication is a substantial gain in information on the life history of these abundant species. Now published: doi.org/10.3390/biology/12070965
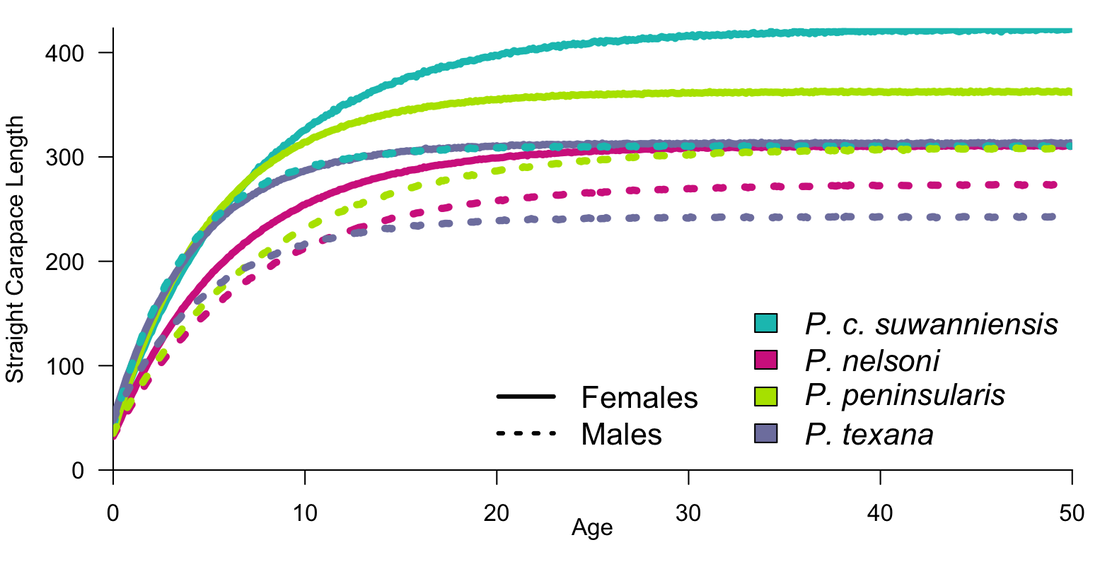
(Above) Predicted length at age for four Pseudemys species (River Cooters) for females and males of each taxa.
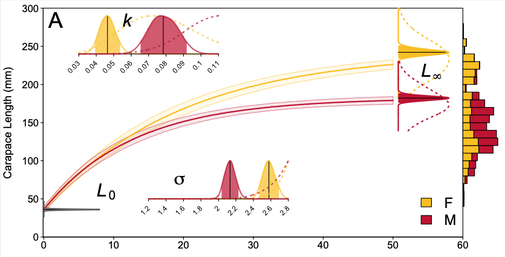 (Above) Predicted carapace length at age for Pseudemys gorzugi (Rio Grande Cooter) for females and males
(Above) Predicted carapace length at age for Pseudemys gorzugi (Rio Grande Cooter) for females and males
Rio Grande Cooters — In collaboration with Dr. Ivana Mali and MJ Suriyamongkol, a multivariate Bayesian von Bertalanffy growth model was estimated using mark-recapture of Rio Grande Cooters (Pseudemys gorzugi) from the Black River in New Mexico, the northern part of the species' range. We found incredibly slow growth in this species but a similar pattern of sexual dimorphism as other Pseudemys, with females far larger than males at their average maximum size. Novel to this model was a joint likelihood modeling the growth in carapace length, plastron length, and body depth while handling the correlations between the different growth parameters. Differences in habitat across the study site led to differences in growth as most individuals had high site fidelity. Now published: doi.org/10.1007/s10750-024-05486-6
This is an active area of research with Dr. Siders collaborating with other research teams to develop advanced Bayesian growth models, synthesize best growth modeling practices, improve age validation analyses, and explore new species-specific applications. Current collaborations are with the Turtle Survival Alliance North American Freshwater Turtle Research Group to develop applications to understudied freshwater turtle species, with NOAA Southwest and Pacific Islands Science Centers to develop latent state models for sea turtles, and with Florida Fish and Wildlife Conservation Commission to apply Bayesian methods to smalltooth sawfish growth.
AQUATIC BIODIVERSITY
The aim of this research is to better understand the processes that influence aquatic community ecology including phylogenetic relatedness, functional similarity, and environmental conditions. While much of this work is ecologically focused, many of the products of this work such phylogenetic and functional relationships between species are useful in the more applied aspects of the lab's research.
Pelagic biodiversity
Pelagic species are often circumglobal, spanning most of the planet's oceans, resulting in pelagic megafaunal communities assembling from recombinations of mostly the same species. Some of the lab's current research explores the assembly dynamics of these combinatorial communities.
|
Shark functional diversity — Initial research was funded by the University of Florida Biodiversity Institute and was focused on dimensions of pelagic shark biodiversity in the North Pacific. For this work we assembled 38 traits from 1225 records from 130 sources, 260 pictographs from seven sources, and 631 teeth photographs from 79 jaw specimens! Using all this trait data, the research identified ten functional groups largely split by life history strategy, habitat, as well as dentition and diet. We found that only bathymetric zone really separated the functional groups of sharks across a suite of macro ecological gradients. Check out the full paper here.
|
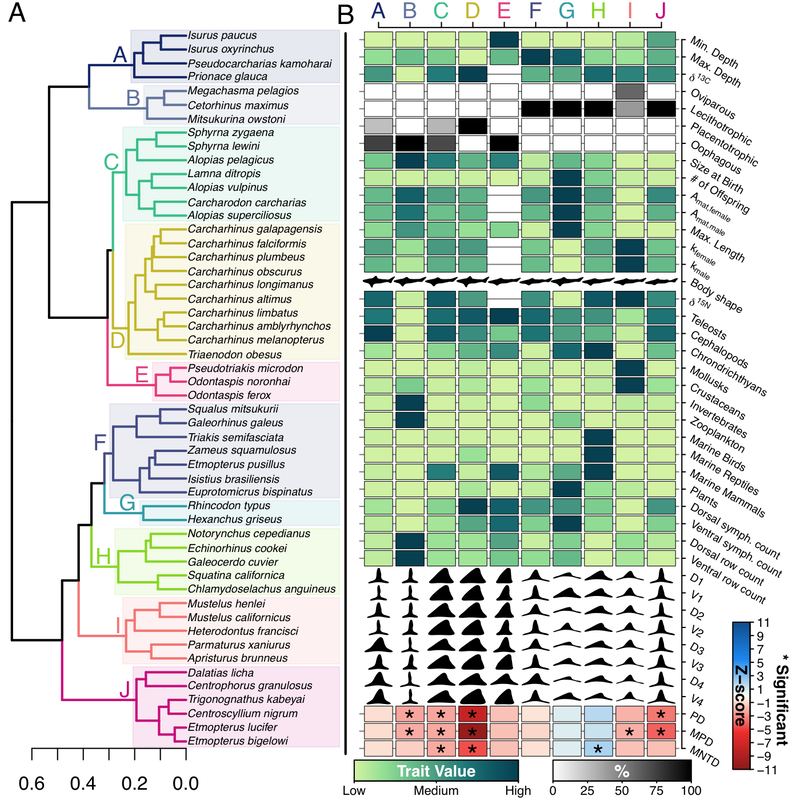
(A) North Pacific shark species and their functional group membership. (B) The mean trait value for each functional group and how clustered it was on the phylogeny.
|
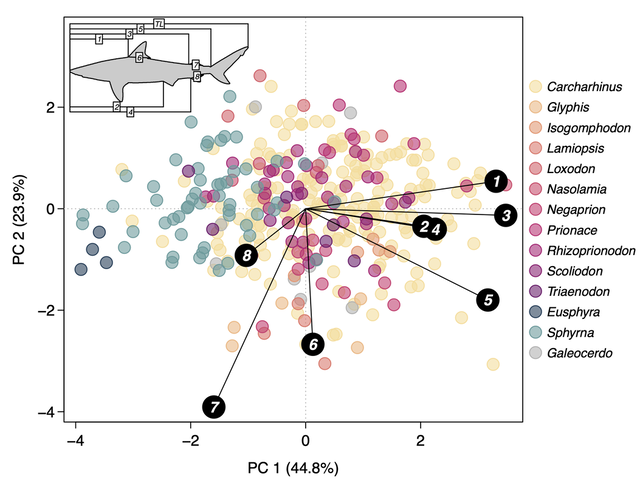 (Above) Principal components analysis of Carcharhinidae and Sphyrnidae proportional length measurements derived from pictographs (scientific illustrations)
(Above) Principal components analysis of Carcharhinidae and Sphyrnidae proportional length measurements derived from pictographs (scientific illustrations)
Scientific illustrations for informing morphological traits — We followed up on our initial research by focusing on developing the use of scientific drawings as morphological traits for ecological and evolutionary analysis. We showed that measurements taken from all the pictographs we collected match those from actual specimens. Most scientific illustrators differ in very minor ways allowing researchers to use most field guides. Lastly, we showed examples of using pictographs with ancestral body shape reconstruction in Lamniformes (mackerel sharks) and phylogenetic generalized linear mixed models in Carcharhinidae & Sphyrnidae (requiem and hammerhead sharks). The former showed the great white/mako shark body shape is relatively new in Lamniformes while the latter showed sharks with narrow species ranges (more endemic) have second dorsal and anal fins as well as smaller heterocercal angles in the caudal fin (dorsal caudal lobe is tilted backwards) . Now published: doi.org/10.1002/ece3.9761
Dimensions of shark biodiversity at depth — In an effort to better understand which sharks interact, we pursued estimating how North Pacific sharks spend their time throughout the water column. To do, we collated 283 existing biotelemetry records from 119 studies with time-at-depth summarizations for 1133 individuals from 35 species. We then trained a hurdle Ensemble Random Forest model to predict time-at-depth for other sharks based on depth ranges reported in field guides and IUCN habitat affinities. From these predictions, we calculated taxonomic, phylogenetic, and functional trait biodiversity metrics and then compared metrics made using just the depth range information versus those made with the extra resolution provided by the time-at-depth information. Not surprisingly, we found that bringing in time-at-depth information resulted in quite different estimates of biodiversity as a function of depth. Now published: doi.org/10.1111/ecog.07249
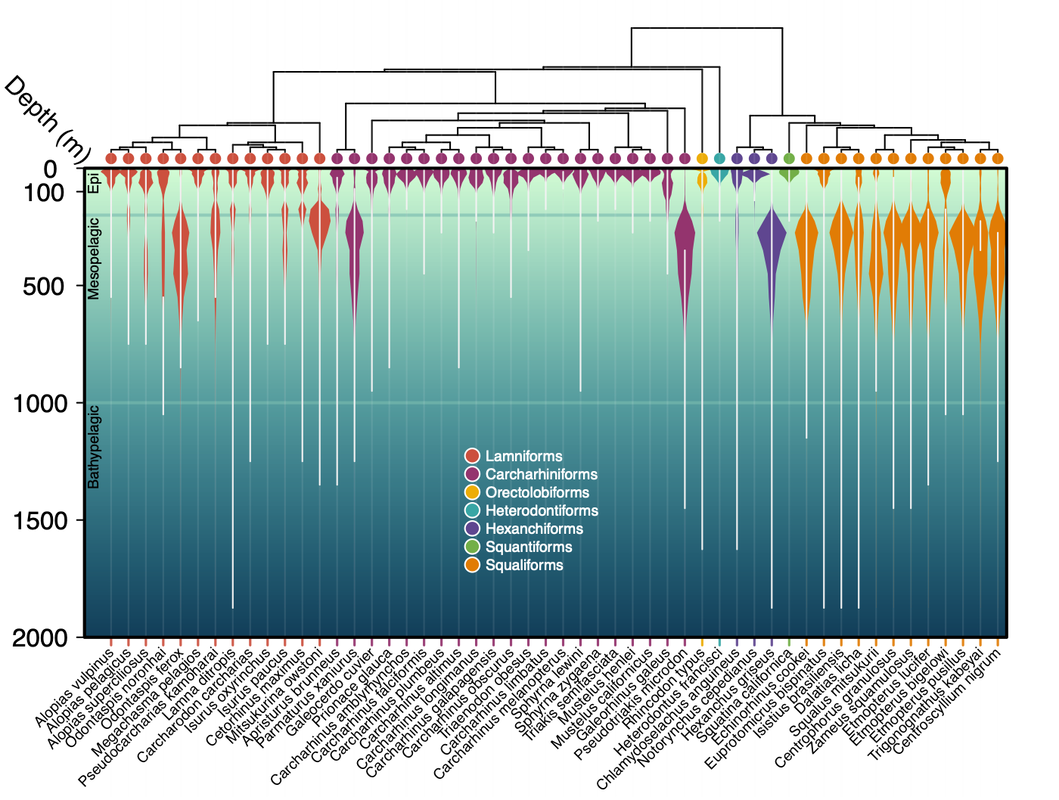
(Above) Predictions of time-at-depth (shaded violins) for North Pacific sharks versus their reported depth range (white line).
This is an active area of research with graduate students working on a variety of research topics and undergraduate researchers working to expand the portfolio of functional traits. An ongoing collaboration with FL SeaGrant and the Return 'Em Right program is working on Gulf of Mexico reef fish applications of the techniques developed for sharks in the North Pacific.
PUBLICATIONS
- Love G.D., Siders Z.A., Gandy D., Pine W.E., Baker S. & Camp E.V. (2024). Estuarine nekton community shows minimal response following large-scale oyster reef habitat loss in Apalachicola Bay, Florida. Hydrobiologia. https://doi.org/10.1007/s10750-024-05548-9
- Long C.A., Ahrens R.N.M., Jones T.T. & Siders Z.A. (2024). A machine learning approach for protected species bycatch estimation. Frontiers in Marine Science 11, 1331292. https://doi.org/10.3389/fmars.2024.1331292
- Siders, Z. A., Trotta, L. B., Patrone, W., Caltabellotta, F. P., Loesser, K. B., & Baiser, B. (2024). Predicting time-at-depth weighted biodiversity patterns for sharks of the North Pacific. Ecography, e07249. https://doi.org/10.1111/ecog.07249
- Siders, Z. A., Murray, C., Puloka, C., Harley, S., Duffy, C., Long, C. A., … Jones, T. T. (2024). Potential of dynamic ocean management strategies for western Pacific leatherback sea turtle bycatch mitigation in New Zealand. Frontiers in Marine Science, 11, 1342475. https://doi.org/10.3389/fmars.2024.1342475
- Suriyamongkol, T., Siders, Z. A., & Mali, I. (2024). Conservation implications of somatic growth and length-at-age in Pseudemys gorzugi, Rio Grande Cooter. Hydrobiologia. https://doi.org/10.1007/s10750-024-05486-6
- Chong, L., Siders, Z. A., Lorenzen, K., Ahrens, R. N. M., & Camp, E. V. (2023). Global synthesis of effects and feedbacks from artificial reefs on socioecological systems in recreational fisheries. Fish and Fisheries, 00, 1–17. https://doi.org/10.1111/faf.12809
- Siders, Z. A., Stratmann, T. A., Turner Tomaszewicz, C. N., Walde, A. D., & Munscher, E. C. (2023). Somatic Growth and Maturity for Four Species of River Cooter Including Pseudemys concinna suwanniensis, P. nelsoni, P. peninsularis, and P. texana. Biology, 12, 19. https://doi.org/10.3390/biology12070965
- Camp E., Siders Z., Ropicki A. & Asche F. (2023). An Introduction to Harvest Tags for Marine Recreational Fisheries. University of Florida, Gainesville, FL. https://doi.org/10.32473/edis-fa253-2023
- Chamberlin D.W., Siders Z.A., Barnett B.K., Ahrens R.N.M. & Patterson W.F. (2023). Highly variable length-at-age in vermilion snapper (Rhomboplites aurorubens) validated via Bayesian analysis of bomb radiocarbon. Fisheries Research264, 106732. https://doi.org/10.1016/j.fishres.2023.106732
- Chamberlin D.W., Siders Z.A., Barnett B.K. & Patterson W.F. (2023). Eye lens-derived Δ14C signatures validate extreme longevity in the deepwater scorpaenid blackbelly rosefish (Helicolenus dactylopterus). Scientific Reports 13, 7438. https://doi.org/10.1038/s41598-023-34680-0
- Chong L., Fisch N., Borsum J.S., Granneman J., Perry D., Love G., et al. (2023). Examining the performance of alternative harvest regulations for short-lived taxa: A case study of Florida Bay scallop management. Fisheries Research 263, 106683. https://doi.org/10.1016/j.fishres.2023.106683
- Siders Z.A., Ahrens R.N.M., Martin S., Camp E.V., Gaos A.R., Wang J.H., et al. (2023). Evaluation of a long-term information tool reveals continued suitability for identifying bycatch hotspots but little effect on fisher location choice. Biological Conservation 279, 109912. https://doi.org/10.1016/j.biocon.2023.109912
- Siders Z.A., Caltabellotta F.P., Loesser K.B., Trotta L.B. & Baiser B. (2023). Using pictographs as traits to explore morphological diversity in sharks. Ecology and Evolution 13. https://doi.org/10.1002/ece3.9761
- Siders Z.A., Martin S.L., Ahrens R.N.M., Littnan C. & Jones T.T. (2023). Update to NOAA Technical Memorandum NMFS-PIFSC-101: Incorporating Uncertainty in Maturation and Latest Fishery Take. Pacific Islands Fisheries Science Center, Honolulu, Hawaii.
- Vilas D., Buszowski J., Sagarese S., Steenbeek J., Siders Z. & Chagaris D. (2023). Evaluating red tide effects on the West Florida Shelf using a spatiotemporal ecosystem modeling framework. Scientific Reports 13, 2541. https://doi.org/10.1038/s41598-023-29327-z
- Vilas D., Fletcher R.J. Jr., Siders Z., Chagaris D. (2022) Understanding the temporal dynamics of estimated environmental niche hypervolumes for marine fish. Ecology and Evolution. doi: 10.1002/ece3.9604.
- Siders Z.A., Westgate A.J., Bell K., Koopman H.N. (2022) Highly variable basking shark (Cetorhinus maximus) diving behavior in the lower Bay of Fundy, Canada. Frontiers in Marine Science. doi: 10.3389/fmars.2022.976857
- Siders Z.A., Trotta L.B., Caltabellotta F.P., Loesser K.B., Baiser B., Ahrens R.N.M. (2022) Functional and phylogenetic diversity of sharks in the northeastern Pacific. Journal of Biogeography. doi: 10.1111/jbi.14383
- Collins A.B., Fluech B., Barbieri L.R., Zangroniz A., Fogg A., Sipos M., Siders Z.A., Camp E.V. (2021). One fish, two fish, twelve years of huge fish: An overview of the Great Goliath Grouper Count in Florida. In: Proceedings of the 74th Gulf and Caribbean Fisheries Institute. Destin, FL.
- Tuckett Q., Lawson K., Lipscomb T.N., Hill J.E., Daniel W., Siders Z.A., (2021) Non-native poeciliids in hot water; the role of thermal springs in facilitating invasion of tropical species. Hydrobiologia. doi: 10.1007/s10750-021-04669-9.
- McCullough J.L.K., Wren J.L.K., Oleson E.M., Allen A.N., Siders Z.A. & Norris E.S. (2021). An Acoustic Survey of Beaked Whales and Kogia spp. in the Mariana Archipelago Using Drifting Recorders. Frontiers in Marine Science. https://doi.org/10.3389/fmars.2021.664292
- Trotta L.B., Siders Z.A., Sessa E.B., Baiser B. (2021) The role of phylogenetic scale in Darwin's naturalization conundrum in the critically imperiled pine rockland ecosystem. Diversity and Distributions. onlinelibrary.wiley.com/doi/10.1111/ddi.13220
- Siders Z.A., Ducharme-Barth N., Carvalho F., Kobayashi D., Martin S., Raynor J., Jones T.T., Ahrens R.N.M. (2020) Ensemble Random Forests as a tool to model rare occurrences. Endangered Species Research. doi.org/10.3354/esr01060
- Siders Z.A., Havens K.E. (2020) Revisiting the Total Maximum Daily Load Total Phosphorous Goal in Lake Okeechobee. Hydrobiologia. doi.org/10.1007/s10750-020-04406-8
- Caltabellotta F.P.†, Siders Z.A.†, Cailliet G.M., Motta F., Gadig O.B.F. (2020) Preliminary age and growth of the deep-water Goblin Shark Mitsukurina owstoni (Jordan, 1898). Marine and Freshwater Research. doi.org/10.1071/MF19370
- Hazelkorn R.A., Wells R.S., Siders Z.A., DeLynn R., Lovewell G.N. (2020) Physical maturity in common bottlenose dolphins (Tursiops truncatus) from Sarasota Bay, FL. Marine Mammal Science. doi.org/10.1111/mms.12733
- Siders Z.A., Allen M.S., Walters C.J., Ahrens R.N.M. (2020) Density-dependent prey behaviors and mutable predator foraging modes induce Allee effects and variability in prey mortality rates. Freshwater Biology. doi:10.1111/fwb.13577
- Rolim F.A.†, Siders Z.A.†, Caltabelotta F.P.†, Rotundo M.M., Vaske-Júnior T. (2020) Growth and derived life history characteristics of the Brazilian electric ray Narcine brasiliensis. Journal of Fish Biology. doi: 10.1111/jfb.14378
- Siders Z.A.†, Martin S.†, Ahrens R.N.M., Jones T.T. Combined effects of U.S. Pacific Islands lonline fisheries on endangered leatherback (western Pacific) and loggerhead (North Pacific) turtle populations. (2020) NOAA Internal Report PIFSC-IR-20-014.
- Martin S.†, Siders Z.A.†, Eguchi T., Langseth B., Yau A., Baker J., Ahrens R.N.M., Jones T.T. (2020) Update to Assessing the Population-level Impacts of North Pacific Loggerhead and Western Pacific Leatherback Turtle Interactions, inclusion of the Hawaii-based Deep-set and American Samoa-based Longline Fisheries. NOAA Technical Memorandum NMFS-PIFSC-101. Honolulu, Hawaii, USA. doi: 10.25923/pnf2-2q77
- Mobini S., Kuliasha C.A., Siders Z.A., Bohmann N., Jamal S-M, Judy J.W., Schmidt C.E., Brennan A.B. (2020) Microtopographical patterns on the surface of neural implants promote different responses in Fibroblasts and Schwann cells. Journal of Biomedical Materials Research Part A. doi:10.1002/jbm.a.37007
- Martin S., Siders Z.A., Eguchi T., Langseth B., Yau A., Baker J., Ahrens R.N.M., Jones T.T. (2020) Assessing the population level impacts of North Pacific Loggerhead and western Pacific Leatherback interactions in the Hawaii-based shallow set fishery. NOAA Technical Memorandum NMFS-PIFSC-95. Honolulu, Hawaii, USA. doi:10.25923/ydp1-f891
- Caltabellotta F.P., Siders Z.A., Murie D.J., Motta F.S., Cailliet G.M., Gadig O.B.F. (2019) Age and growth of three endemic threatened guitarfishes (Pseudobatos horkelii, Pseudobatos percellens and Zapteryx brevirostris) in the Western South Atlantic (Chondrichthyes, Rhinopristiformes). Journal of Fish Biology. doi: 10.1111/jfb.14123
- Matthias B.G., Ahrens R.N.M., Allen M.S., Tuten T., Siders Z.A., Wilson K.L. (2018) Understanding the effects of density and environmental variability on the process of fish growth. Fisheries Research. 198. 209-219. doi: 10.1016/j.fishres.2017.08.018
- Lynch A.J., Cooke S.J., …, Siders Z.A., Taylor W.W., Youn S. (2017) Grand challenges in the management and conservation of North American inland fish and fisheries. Fisheries. 42(2): 115-124. doi: 10.1080/03632415.2017.1259945
- Koopman H.N., Westgate A.J., Siders Z.A. (2015) Declining fecundity and factors affecting embryo quality in the American lobster (Homarus americanus) from the Bay of Fundy. CJAFS. 72: 353-363. doi: 10.1139/cjfas-2014-0277
- Koopman H.N., Westgate A.W., Siders Z.A., Cahoon L.B. (2014) Rapid sub-surface ocean warming in the Bay of Fundy as measured by free-swimming basking sharks. Oceanography 2: 14-16. doi: 10.5670/oceanog.2014.32
- Westgate A.W., Koopman H.N., Siders Z.A., Wong S.N.P., Ronconi R.A. (2014) Population density and abundance of basking sharks in the lower Bay of Fundy, Canada. Endangered Species Research. 23: 177-185. doi:10.3354/esr00567
- Siders Z.A., Westgate A.J., Johnston D.J., Murison L.M., Koopman H.N. (2013) Seasonal variation in the spatial distribution of basking sharks (Cetorhinus maximus) in the lower Bay of Fundy, Canada. PLoS ONE 8(12): e82074. doi:10.1371/journal.pone.0082074
- Koopman H.N., Siders Z.A. (2013) Variations in egg quality in blue crabs, Callinectes sapidus, from North Carolina: does female size matter? Journal of Crustacean Biology. 33. 481-487. doi: 10.1163/1937240X-00002152